I'll post this now as I've gone off down a related but more concise rabbit hole for the next posts.
Okay. Here are a couple more building blocks for the future, snippets from the slightly dubious
Here is the pattern of ROS generation from isolated mitochondria fed with increasing concentrations of palmitoyl carnitine:
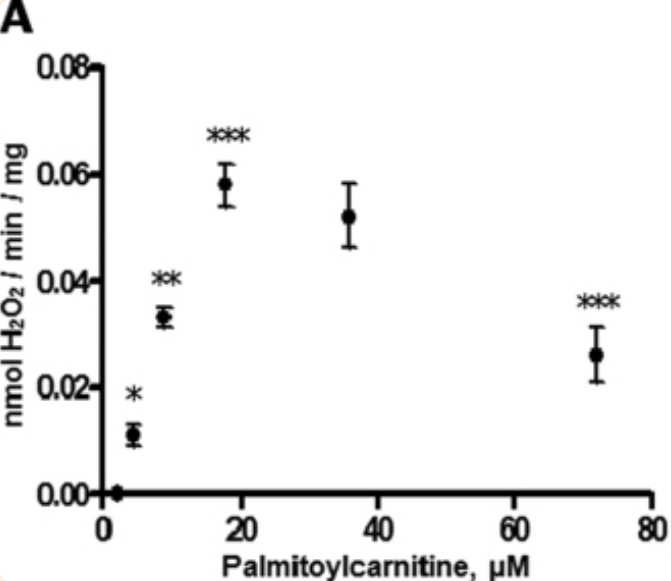
Clearly there is a rise in ROS with increasing PCarn exposure, peaking at 18microM and decreasing to 72microM. There were too few functional preparations to include the 36microM value in statistical analysis. There is also evidence of uncoupling at 72microM. So 18microM gives maximum ROS generation. All measurements were taken under oligomycin to mimic state 4 respiration, ie fed with PCarn but conversion of ADP to ATP was not allowed.
And here is the delta psi developed by PCarn (plus extra carnitine) at increasing concentrations of PCarn. Note that there is a consistent delta psi estimated using between 3microM and 18microM PCarn with no suggestion of an upward trend:

Other places in the paper suggest that this delta psi is around 140mV (see below) and that exposure to PCarn at lower or higher concentrations will produce less ROS than the 18microM exposure does. To get more information about ROS generation they used oligomycin to establish a high delta psi then lowered it a little with 15pM FCCP (right hand end of the graph below) or a lot with 100pM FCCP (left hand end of the graph. As 18microM PCarn can only support 140mV I'm guessing that both voltages are below this value.

Here PCarn, the triangles, shows significant ROS generation at low delta psi with a modest rise to high delta psi, the latter being a little under 140mV. Glutamate shows a delta psi responsive rate of ROS generation.
In this final graph, ignoring the O2 consumption, we can look at the end derived delta psi values from three substrates:

The (expected) delta psi for 18microM PCarn is at 140mV (this is the max which PCarn at any concentration will support in this model), for saturating glutamate with blocked complex II it's 170mV and for succinate (driving through complex II) it's somewhere around 190mV.
What I am driving at is that palmitate, whilst generating ROS, does this without the requirement for, and not as a result of, high delta psi. For glutamate the higher the delta psi, the higher the ROS.
There is no suggestion that PCarn at 18microM is uncoupling yet delta psi is relatively low. What interests me is whether this is being achieved through the inhibition of complex I. In the last post I talked about the role of glutathiolation of proteins as a consequence of localised ROS generation and the role which that glutathiolation might have in the avoidance of situations where unbridled supply of metabolic substrate would lead to very high delta psi and an unacceptable rise in ROS generation. Recall Nick Lane's concept that a cell will sacrifice ATP production from ox-phos rather than generate excess ROS.
How much glutathione was available and what state complex I's glutathiolation might be were not in the remit of this study. So a lot of unknowns.
These are isolated mitochondria. There are, in real life, many controls to the supply of calories to mitochondria, as near by as the carnitine palmitoyltransferases, further away at the cell surface (GLUT4 and CD36) and as far away as adipocytes and hepatocytes.
Again I have further ideas about ROS and it has to be made very clear that I am making huge extrapolations from isolated mitochondria towards what might happen in an intact cell with insulin exposure, insulin receptors, phosphatases, a controllable supply of Ca2+, superoxide dismutase, catalase etc etc, all in the correct locations to do what is impossible for an isolated mitochondrion to do.
So caution. But ultimately ROS from fatty acid oxidation generates ROS without the need for a high delta psi. Primarily NADH supplying substrates produce ROS in association with (and probably caused by) high delta psi.
Peter

3 comments:
I try to keep up, but it's difficult: high delta psi means energy depleted cell correct?
So fatty acid oxidation generates ROS, even without a high delta psi? Is that generally speaking, or only in this experiment?
And what would that mean in real life, more uncoupling?
Hi Stefan, high delta psi here is an energy depleted cell/mitochondrion because it is being bought at the cost of oligomycin, an antibiotic which blocks ATP synthesis. IRL an high delta psi is the result of calories entering the cell in excess of the cell's needs, ie energy replete. ATP exerts negative feedback on ATP synthase so raises delta psi giving more ROS and disabling insulin signalling. Fatty acids generate ROS w/o high delta psi, they'll disable insulin signalling just because the cell has fatty acids so doesn't need insulin facilitated glucose uptake. Both ROS sources inhibit glucose uptake but FFAs do it w/o high delta psi. My opinion is that high delta psi does damage with its ROS, ie they are an emergency response. FFA oxidation is planned ROS, they'll do Good Things. Watch my biases.
Oh, and yes, uncoupling is the response to ROS, seems to happen with both sources and is a potent technique to suppress insulin signalling.
Peter
Peter, that was a very good summary in your reply to Stefan. I have followed you for years, but some of the details of core mechanism keep slipping my mind. Could you try to write a reference / primer kind of post that will be the go to place for us to keep up? Much like your reply above but maybe 10x the length, so a little more detail?
Thanks
Post a Comment