The CoQH2/CoQ Ratio Serves as a Sensor of Respiratory Chain Efficiency.
We should all have a picture of the ETC looking a bit like this, omitting mtG3Pdh and ignoring supercomplex formation:

Guarás et al set up a model, a proof of principle extreme. They blocked the ETC completely at either complex III, or at cytochrome C or at complex IV. They even discussed short term oxygen deprivation as a generator of ROS, equivalent of blocking the extreme end point of the ETC. If you completely block the ETC in this way then any input through FADH2 containing enzymes to CoQ will always generate a massively reduced CoQ pool, one prerequisite for reverse electron transport (RET),
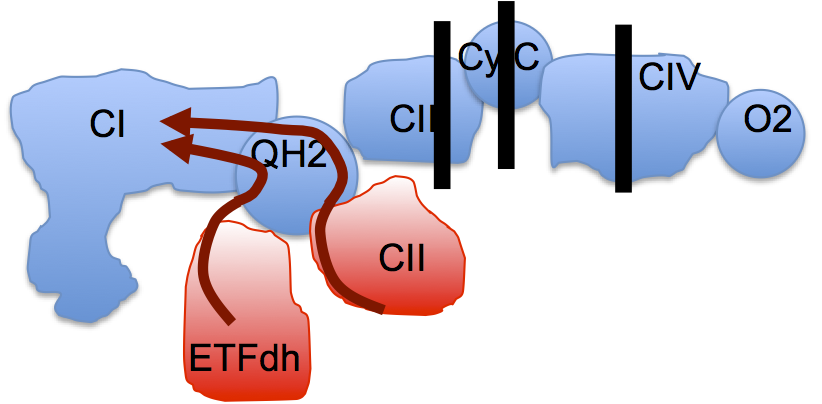
leading to the near complete disassembly of complex I. It's a model, an extreme version of reality. Fascinating in its own right, as was the ability of the fungal CoQH2 oxidase (AOX) to protect complex I completely from any of the engineered ETC defects:

AOX may not pump any protons but it does preserve complex I by reducing the extreme levels of CoQH2 which drive RET.
They subsequently went on to look at more physiological ways to generate RET and came to the conclusion that, as the balance of inputs from NADH vs FADH2 shifted, then the amount of complex I relative to complex III would need to be altered. RET is the physiological signal to balance complexes I and III against NADH and FADH2 supply.
Elegant is not the word.
They even went on to ascertain which cysteine residues were preferentially oxidised to disassemble the complex. They group around the FMN and the CoQ docking area, surprise surprise... OK. If you insist, here is the ribbon diagram from fig 5:

There's a lot of explanation in the text of what the colours and the asterisks mean.
All I really wanted to lay down with this post is that there is a physiological process where FADH2 inputs control complex I abundance. That is how it should be.
When you want a pathological model of complex I destruction, pathological levels of FADH2 input will deliver.
Peter
